The Approach
Three complementary approaches guide our work.
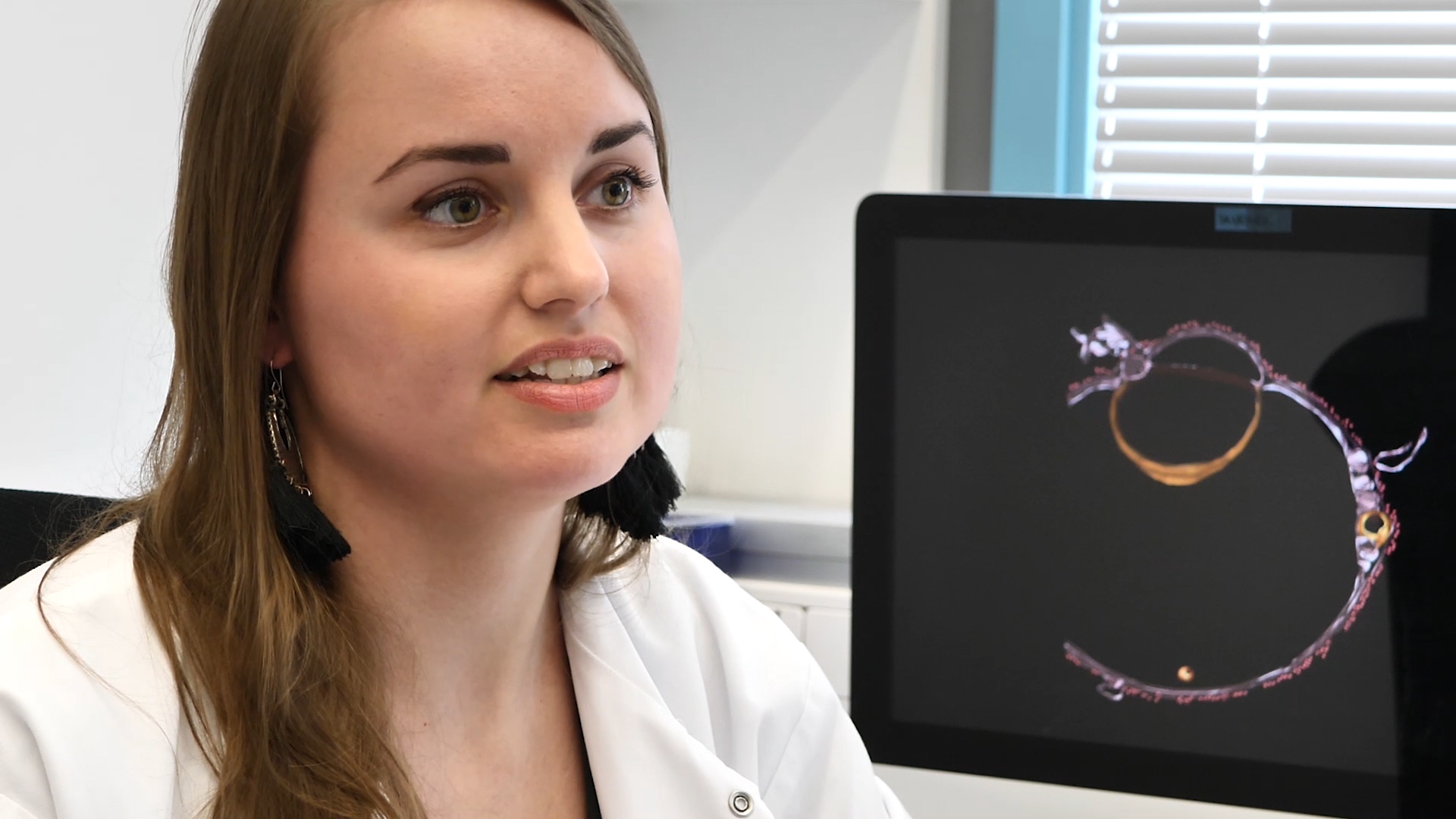
First, we study how the nuclear envelope integrates lipid metabolism with nuclear pore complex biogenesis. Using biochemical reconstitution and structural approaches, we dissect how nuclear pores assemble in membranes and how lipid metabolism at the inner nuclear membrane shapes nuclear architecture, including the formation of nuclear lipid droplets.
Second, we investigate how biomolecular condensates organize transcription and genome folding. Combining quantitative single-molecule techniques, chromosome conformation capture methods and genetics, we directly probe how condensates interact with chromatin and the transcription machinery.
Third, we explore how cells preserve functional architecture under extreme physiological stress using tardigrades (water bears) as a model system. Using cryo-electron tomography, quantitative proteomics and cellular biophysics, we examine how macromolecular systems remain functional during desiccation and other extreme conditions.
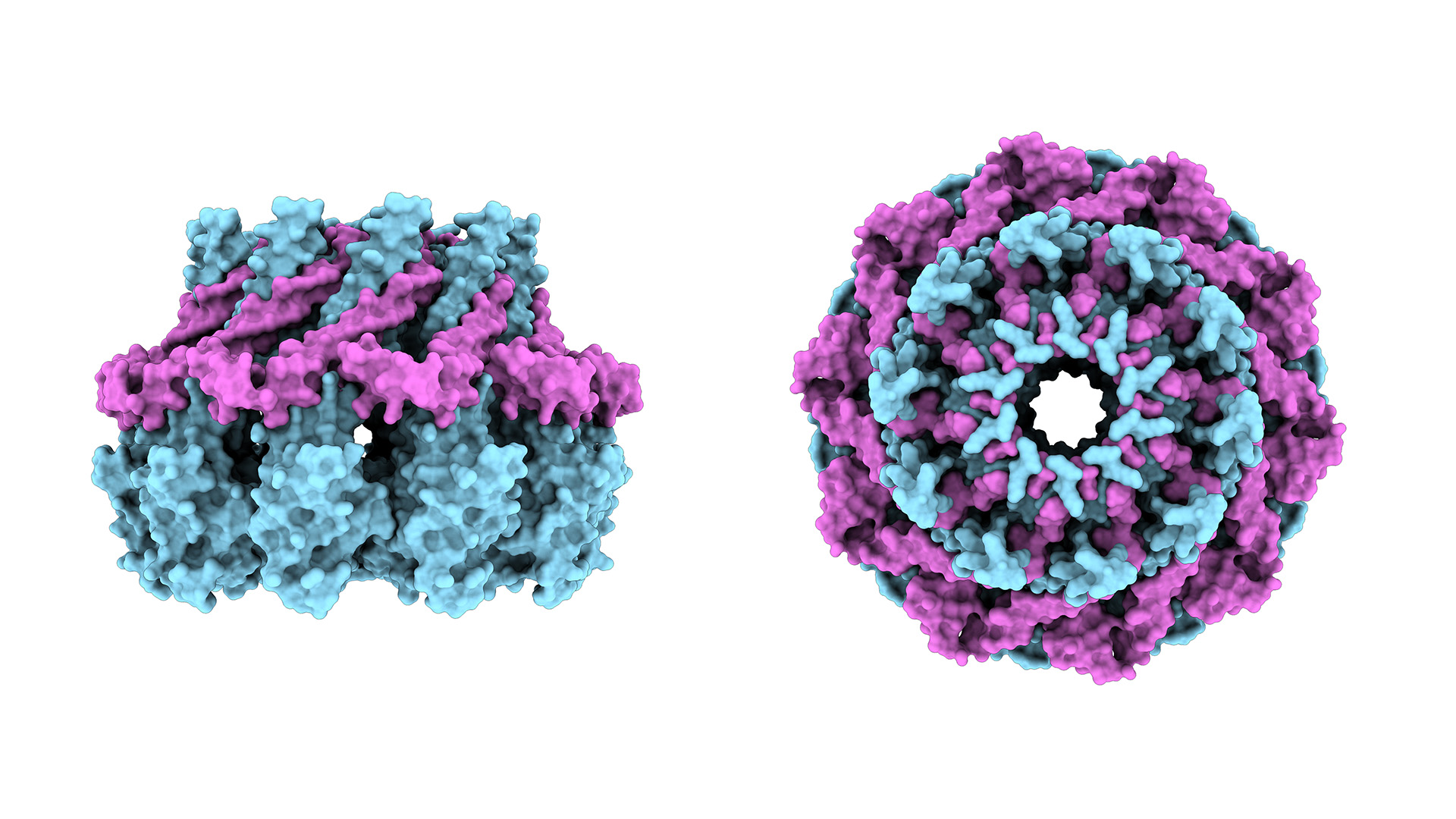
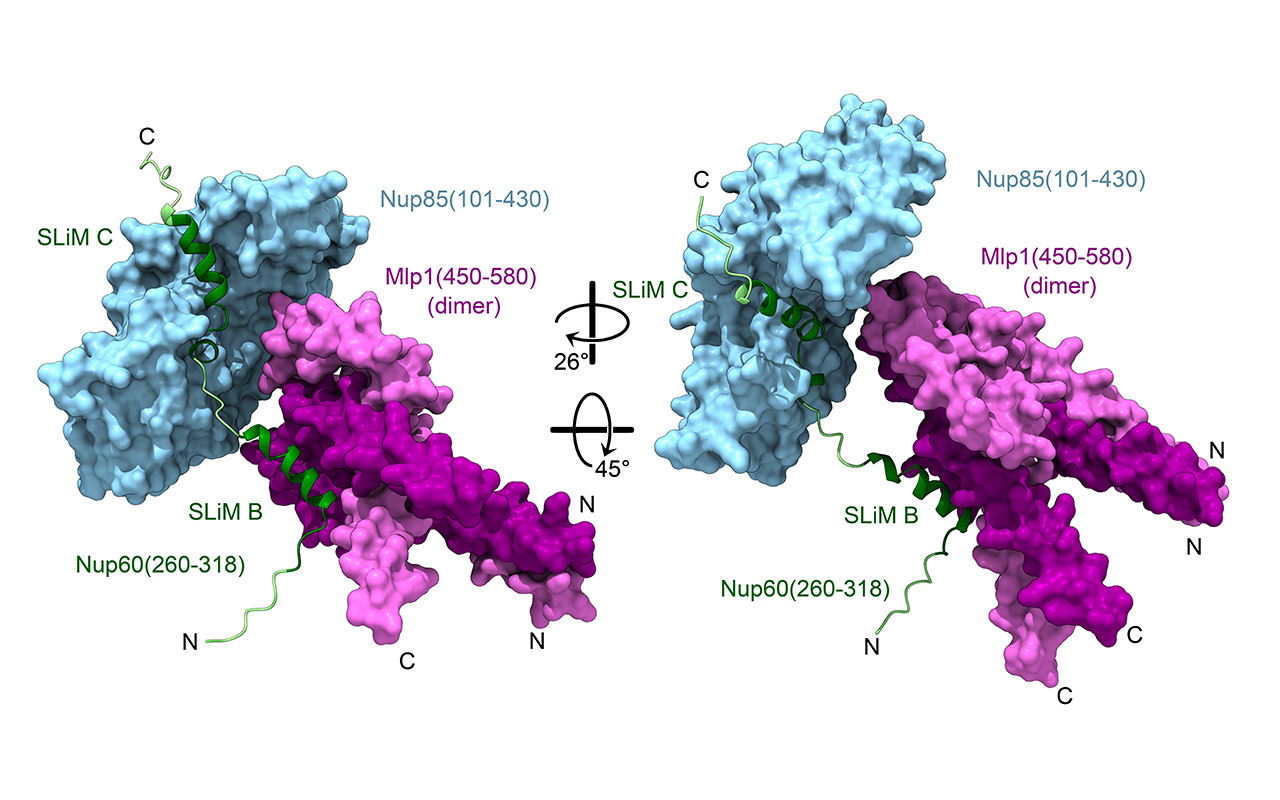
 Alwin Köhler, Max Perutz Labs Balanced lipid saturation is essential for nuclear envelope and nuclear pore complex architecture and function. Romanauska and Köhler identify mechanisms that prevent a cell nucleus from becoming edgy, as seen in the EM-based 3D reconstruction of the nuclear envelope (gold) and endoplasmic reticulum (blue).")


")


















 Laura Gallego")